Lexikon der Biologie: Vogelzug
ESSAY
Peter Berthold
Vogelzug
Vogelzug bezeichnet i.e.S. die regelmäßigen jahresperiodischen (saisonalen) Pendelbewegungen zwischen einem Brutgebiet (Brutrevier) und einem Ruheziel („Winterquartier“, „Ruhequartier“) von Vögeln. Man spricht auch von Jahreszug, da die Wanderungen (Tierwanderungen) jährlich stattfinden. Der Vogelzug besteht aus Herbstzug und Frühjahrszug, sollte aber besser neutral als Wegzug und Heimzug bezeichnet werden, da er – weltweit gesehen – ganzjährig stattfindet. I.w.S. versteht man unter Vogelzug auch weniger regelmäßige Wanderungen wie Invasionen, Nomadenwanderungen, Ausbreitungsbewegungen (Ausbreitung), Folge- und Fluchtbewegungen, Teilzug (s.u.) sowie Dispersionswanderungen (Dismigration: Zerstreuungswanderungen, vor allem von Jungvögeln, zur Verteilung über das engere Brutgebiet hinaus) und Abmigration (das sog. Auswandern, vor allem bei Enten, bei dem nach Verpaarung im Ruheziel ein Partner einen anderen in ein zum Teil weit entlegenes Brutgebiet „entführt“). Im Extrem kann man alle Vogelbewegungen von einer räumlichen Einheit zu einer anderen, die über die täglichen Routinebewegungen (zur Nahrungssuche [Nahrungserwerb], zum Schlafplatz usw.) hinausgehen, einschließlich Verdriftung (Drift), z.B. von Irrgästen (Alieni), als Vogelzug bezeichnen.
Geschichte der Zugforschung
Die Anfänge der Vogelzugforschung sind unbekannt. Der Beginn der wissenschaftlichen Betrachtung wird Aristoteles zugeschrieben, der noch an den „Winterschlaf“ von Zugvögeln, vor allem bei Schwalben, sowie an die sog. Transmutationstheorie (Umwandlung von Winter- in Sommervögel und umgekehrt) glaubte. Nachdem es sogar Vorstellungen über Vogelwanderungen bis zum Mond gab, zeigte vor allem Friedrich II. von Hohenstaufen (1194–1250) erste realistische Einschätzungen über den Vogelzug. Einige Jahrhunderte später schrieb der Verhaltensforscher Baron Ferdinand Adam von Pernau (1660–1731) über die Steuerungsgrundlagen des Vogelzugs, und gegen 1800 erfolgte dann die Entdeckung der Zugunruhe (Ausdruck eines endogenen Zugtriebs) durch Johann Andreas Naumann (1747–1826; Naumann [J.F.]). Weitere Überlegungen zur endogenen Kontrolle des Vogelzugs stellten Hermann Samuel Reimarus (1760) und Legg (1780) vor. Auf dem ersten Internationalen Ornithologen-Kongreß (1884) in Wien erfolgte der Beschluß zur Einrichtung von Beobachtungsstationen zur systematischen Erforschung der Wanderungen, die 1901 zur Gründung einer ersten Vogelwarte (im Sinne einer Sternwarte) führte (zuerst „Vogelwarte Rossitten“ in Ostpreußen, heute „Vogelwarte Radolfzell“ am Bodensee, Forschungsstelle der Max-Planck-Gesellschaft). In der „Vogelwarte Rossitten“ erfolgte die Institutionalisierung einer zuvor privat entwickelten Methode: der Beringung (individuelle Kennzeichnung mit Metallringen am Bein). Nach der „Periode der Beobachtungen“ folgte ab 1925 die „Periode der experimentellen Vogelzugforschung“ mit ersten Untersuchungen u.a. zum Einfluß von Photoperiode (Photoperiodismus) und Hormonen, zur Physiologie des Zugtriebs und zur Orientierung (durch Versetzungs- und Auflaßversuche).
Evolution des Vogelzugs
Der wesentliche Schrittmacher für Vogelwanderungen wird heute allgemein in Nahrungsgrundlagen gesehen. Sie wirken zumeist als ultimate (mittelbare, evolutiv selektionierende) Umweltfaktoren (ökologische Faktoren, Umwelt) auf zweierlei Weise: zum einen führt die jahreszeitliche Nahrungsverknappung (z.B. Rückgang von Insekten in höheren geographischen Breiten im Winterhalbjahr) zum saisonalen Wegzug von Arten (etwa von Insektenfressern wie Schwalben), zum anderen werden Gebiete mit jahreszeitlich besonders günstigen Bedingungen zum Brüten aufgesucht, wie z.B. die Tundra mit extrem langen Tagen im Sommer, reichem Nahrungsangebot, geringem Feinddruck (Feindfaktor) usw. von Gänsen, Schnepfenvögeln und anderen. Von den proximaten (unmittelbaren, aktuell verhaltensregulierenden) Umweltfaktoren spielen für den Vogelzug in erster Linie die Photoperiode (jahreszeitliche Änderungen der Tageslichtdauer; Lichtfaktor) sowie auch Wetterfaktoren (Klima) und Populationsdichte (Abundanz) eine große Rolle (s.u.). Früher nahm man an, daß der Vogelzug in Folge der Eiszeiten (Pleistozän; Paläoklimatologie) oder im Zuge der Kontinentalverschiebung (Kontinentaldrifttheorie) entstanden sei. In den letzten Jahrzehnten verdichtete sich jedoch die Ansicht, daß der Vogelzug sehr alt ist – möglicherweise (fast) so alt wie die Vögel selbst. Er soll bereits in den Tropen, oder zumindest unter tropisch-subtropischen Bedingungen, unter denen Vögel evoluiert sind und ursprünglich gelebt haben, entstanden und durch einen „Verhaltenssprung“ (Mutation) aus Standvögeln hervorgegangen sein – vermutlich polyphyletischen Ursprungs, also mehrfach unabhängig voneinander. Die neueste Theorie zur Evolution, Steuerung und Anpassungsfähigkeit des Vogelzugs (Berthold 1999) postuliert: Die weitaus häufigste Lebensform der heute lebenden Vogelarten ist die des Teilziehers, wobei ein Teil einer Population im Brutgebiet verbleibt, ein anderer Teil regelmäßig wegzieht (wie z.B. in Mitteleuropa bei den meisten Amselpopulationen [Amsel]). Teilzug kommt bei Vögeln in allen Kontinenten und systematischen Gruppen vor und ist zudem eine ursprüngliche, entwicklungsgeschichtlich sehr alte Verhaltensweise, die in allen Wirbeltierklassen, bei sehr vielen Nichtwirbeltieren und sogar bei Pflanzen auftritt. Es ist wahrscheinlich, daß Vögel die Fähigkeit zum Teilzug bereits von Vogelvorfahren geerbt haben und damit allgemein mit Erbanlagen (Gen) für Ziehen und Nichtziehen ausgestattet sind. Das Verhältnis der beiden Gentypen ist entscheidend dafür, ob Vogelindividuen eher Zugvögel oder Standvögel sind. Durch Selektion können sich in Teilzieherpopulationen die Anteile von Zug- und Standvögeln innerhalb weniger Generationen stark verschieben. Im Experiment ließen sich aus Teilziehern – Mönchsgrasmücken aus Südfrankreich – in 3–6 Generationen reine Zug- bzw. Standvögel züchten. Damit können Vogelpopulationen ihr Zug- oder Standvogelverhalten schnell an neue Umweltbedingungen anpassen (Anpassung, genetische Flexibilität), wie z.B. derzeit an Verhältnisse, die durch die rezente Klimaerwärmung bedingt sind (s.u.). Im Extremfall können Teilzieher so stark in eine Richtung selektiert werden, daß Populationen phänotypisch (Phänotyp) als reine Zug- oder Standvögel erscheinen, obwohl sie im Genom oder zumindest im Genpool auch noch Gene der anderen Verhaltensweise besitzen. Der genotypische [Genotyp] oder potentielle Teilzug ist also ein quantitativ-genetisches Merkmal.
Untersuchungsmethoden
Anfänglich waren nur Sichtbeobachtungen (Nachtzug vor allem vor der Mondscheibe) von Bedeutung, später erfolgten zunehmend Untersuchungen, die auf Jagd und Sammeltätigkeit aufbauten. Ab Ende des 19. Jahrhunderts kamen dann moderne Verfahren individueller Kennzeichnung (Beringung) hinzu, die ab 1903 in der „Vogelwarte Rossitten“ in großem Stil ausgebaut wurden. Die Methode der Beringung ermöglichte über Wiederfunde (geschossener oder tot gefundener Vögel) u.a. die Rekonstruktion von Wanderwegen, Zugzeiten, Ruhezielen und wird heute in Deutschland von 3 Vogelwarten (Helgoland, Radolfzell, Hiddensee) organisiert. Bisher wurden weltweit über 200 Millionen Beringungen durchgeführt, die das erdumspannende Streckennetz von Zugrouten weitgehend aufklärten. Nach dem Zweiten Weltkrieg wurde die Radarornithologie, die vor allem den Nachtzug „sichtbar“ macht, maßgeblich ausgebaut. Hierdurch erfolgt heute die zum Teil kontinentweite Erfassung von Zugvogelbewegungen (in den USA, mit Überwachungsradar). Zielverfolgungsradar und Telemetrie „besenderter Individuen“ ermöglichen die Erfassung von Einzelzugabläufen, mit terrestrischer Telemetrie über kurze Distanzen, mit Satelliten-Telemetrie unbegrenzt. Letztere eröffnete in der Ornithologie und vor allem in der Zugforschung eine neue Dimension. Obwohl erst 1984 in den USA und 1990 in Europa begonnen, wurden inzwischen bereits über 100 Großvogelarten untersucht – derzeit minimal bis zu etwa Taubengröße möglich. Bei Störchen wurden die Untersuchungen bereits bis zu etwa 5 Jahren lückenlos durchgeführt, mit zum Teil über 20 Ortungen pro Tag und mehr als 2000 Ortungen pro Saison. Näherrückendes Fernziel ist die Erfassung von Lebenswanderstrecken und -verhalten langlebiger Arten. Standardisierte Fangverfahren (auf Fangstationen, zum Teil über Jahrzehnte hinweg) vermitteln in Verbindung mit der Beringung Weg- und Durchzugsmuster, Analysen der Rastbiologie und ökophysiologischer (Ökophysiologie) Vorgänge, wie z.B. der Fettdeposition in Bezug zur Rastplatzqualität, Veränderungen in Abhängigkeit von der globalen Klimaerwärmung usw. Eine Schlüsselrolle kommt der systematischen Erfassung der Zugunruhe (s.o.) zu. Diese nächtliche Aktivität (hauptsächlich „Schwirren“, „Ziehen im Sitzen“: Flügelschlagen mit hoher Frequenz und geringer Amplitude; vgl. Abb. 1 ) gekäfigter Vögel normalerweise tagaktiver, aber nachts ziehender Arten (s.u.) gibt ein erstaunlich gutes Abbild des Zugs in freier Natur wieder. Unter anderem können Zugbeginn, -ende, -dauer und –intensität untersucht werden. In Verbindung mit Selektions- und Kreuzungsversuchen (Kreuzung) gelang es über Zugunruhemessungen (hauptsächlich an Grasmücken und Rotschwänzen; vgl. Abb. 2 ) in der Vogelwarte Radolfzell, neue Arbeitsgebiete zu eröffnen: die Genetik und die experimentelle Evolutionsbiologie des Vogelzugs, mit denen genetische Grundlagen des Zugs erforscht werden (s.u.). Eine Reihe von Orientierungskäfig-Typen ermöglicht die Bestimmung programmierter Zugrichtungen ( vgl. Abb. 5 ). Für zunehmend neue Fragestellungen kommen immer mehr moderne Methoden der Biologie, Medizin, Physik usw. zum Einsatz, wie z.B. die molekulare Genetik, die Kernspintomographie oder die Untersuchung stabiler Isotope.
Umfang und Erscheinungsformen
Mehr als die Hälfte der heute lebenden rund 10.000 Vogelarten führt regelmäßig Wanderungen durch. Die Größenordnung jährlich ziehender Individuen beträgt ca. 50 Milliarden. Das Zugstreckennetz umspannt die ganze Erde, wobei Zugvögel sowohl Kontinente als auch Ozeane und Eisfelder überqueren einschließlich der Wüsten, Regenwälder und Hochgebirge. Vogelzug kennt somit letztlich keine „ökologischen Barrieren“. Die Zugstrecken variieren von wenigen hundert Metern (bei Vertikalzug, in Gebirgen) bis etwa zur Hälfte des Erdumfangs (bei Küsten-Seeschwalben: Weg- und Heimzug zusammen bis zu 50.000 km, Lebenswanderstrecke mehr als 1 Million km). Non-Stop-Flüge reichen bei Schnepfenvögeln bis zu 10.000 km und bei Kleinvögeln bis ca. 3000 km. Man unterscheidet Kurzstreckenzieher (unter den europäischen Vögeln z.B. Kiebitz, Bleßhuhn, Graureiher und von den Singvögeln Wacholder-Drossel, Wiesen-Pieper, Heide- und Feld-Lerche), Mittelstreckenzieher (z.B. Amsel, Rotkehlchen, Haus-Rotschwanz) und Langstreckenzieher (u.a. Seeschwalben, Wachtel, Kuckuck, Mauer-Segler, Wiede-Hopf und Singvögel, wie Schwalben, Grasmücken, Gartenrotschwanz oder Nachtigall; vgl. Abb. 3 ). Langstreckenzieher sind weitgehend Interkontinentalzieher, die von Europa vor allem bis ins südliche Afrika ziehen.
Die früher verwendete Bezeichnung Strichvögel ist unzureichend definiert und sollte nicht mehr verwendet werden. Der Hauptzug verläuft in Höhen von 50–3000 m; Hochgebirge werden in bis zu 10.000 m Höhe überquert (gemessener Rekord: 11.300 m). Vielerorts – wie in Mitteleuropa – dominiert der Nachtzug (bei vielen Langstreckenziehern, vor allem Insektenfressern, Entenvögeln, Schnepfenvögeln), der der Zeitminimierung dient. Tagzug kommt vor allem bei segelfliegenden, im Gleitflug wandernden Arten, wie Störchen und Greifvögeln, vor. Häufigste Form ist der Breitfrontzug (querfeldein über Land, Wasser, Gebirge usw.) im Gegensatz zum Schmalfrontzug (in Zugkorridoren oder Zugstraßen, oft Leitlinien wie Flußtälern folgend; z.B. bei verschiedenen Gänsen, Entenvögeln und auch Singvögeln). Ein Schmalfrontzug kann zu sog. Zugscheiden führen. Hierbei erfolgt eine Trennung nach Populationen, wie z.B. beim Weißstorch in „Ost-“- und „West“-Störche, die über Israel bzw. Gibraltar nach Afrika wandern. Viele weitere Unterscheidungen betreffen die räumliche Verteilung (z.B. Trichter-, Fächer-, Ketten-, Überspringzug, differenziertes Zugverhalten nach Alter und/oder Geschlecht), die ökologische Ausrichtung (u.a. Mauserzug, Zwischenzug, Umkehrzug, Folge- und Fluchtbewegungen, Nomadisieren, zum Teil in Zwischenziele). Beim Teilzug unterscheidet man die regelmäßige Form des obligaten (s.o.) vom fakultativen Teilzug, der unregelmäßige Invasionen (Massenwanderung) bewirkt. Die Hauptfortbewegung auf dem Zug erfolgt durch Fliegen (Flug), in geringem Umfang auch durch Schwimmen und Laufen. Die Hauptwegzugrichtungen sind äquatorwärts gerichtet; in der Nordhemisphäre durch Süd-, in der Südhemisphäre mit Nordkomponenten (sog. Australzug) gekennzeichnet. In Nordamerika überwiegen die Nord-Süd-Zugrichtungen, in Eurasien (Europa, Asien) mehr die Ost- und Westabweichungen wegen der ost-west-ausgerichteten Gebirge (Alpen, Himalaya). Viele Arten wandern geradlinig, andere mit systematischen Richtungsänderungen (Schleifen-, Bogenflug, Zugknicks). Brutgebiete und „Winterquartiere“ werden mit großer Präzision immer wieder aufgesucht (Ortstreue). Ziehen an sich stellt für die Vögel kein erhöhtes Risiko dar. Viele Zugvogelarten kommen mit 1 Jahresbrut aus (im Gegensatz zu Überwinterern in höheren Breiten mit oftmals mehreren Bruten). Die Aufenthaltsdauer im Brutgebiet ist sehr variabel – von fast 12 Monaten (bei Wetterflüchtern) bis zu 11/2 Monaten (bei extremen Langstreckenziehern, z.B. Sumpf-Rohrsänger).
Anpassungen und physiologische Grundlagen
Vögel sind für ihre umfangreichen Wanderungen wie keine andere Tiergruppe (Tiere) durch ihr Flugvermögen (Flugmechanik) und damit einhergehende Merkmale präadaptiert (Präadaptation). Hierzu gehören der leichte Körperbau (Pneumatisation der Knochen; pneumatische Knochen), ein besonders effizientes Herz-Kreislauf- und Atmungssystem (Parabronchiallunge als „Durchströmlunge“ [Lungenpfeifen, Atmungsorgane III ], Hämoglobin-Polymorphismus mit Hämoglobintypen [Hämoglobine] für verschiedene Höhenbereiche) und zahlreiche Spezialanpassungen der Muskeln (große Brustmuskeln als Hauptflugmotoren, mit bis zu 35% des Körpergewichts, schnelle Muskelfasern mit hoher Dichte an Blutkapillaren, Mitochondrien und Myoglobin; Flugmuskeln). Langstreckenzieher besitzen besonders lange und vor allem spitze Flügel (Vogelflügel), die zwar die Manövrierfähigkeit beeinträchtigen, aber aufgrund günstiger Luftwirbelbildung für Streckenflüge besonders effizient sind. Ausgeprägte Zugvögel erreichen Zug-Bereitschaft in der sog. Zugdisposition, einem komplexen verhaltensphysiologischen Zustand, der vor allem folgendes umfaßt: a) vielfach (zum Teil schon im Ei) beschleunigte Jugendentwicklung für deren Abschluß vor dem Aufbruch, b) meist schon vor Zugbeginn einsetzende Hyperphagie (verstärkte Nahrungsaufnahme) zur Bildung von Fettdepots (als Treibstoff, oft verbunden mit Verdoppelung des Körpergewichts; Depotfett, Fettspeicherung), c) Umstellung auf Fettstoffwechsel und Reduktion der Kohlenhydratreserven (Kohlenhydrate), d) zum Teil Ballastreduktion durch Dehydrierung und während des Zugs durch temporäre Rückbildung des Magen-Darm-Trakts und anderer Organe; e) vielfach erfolgt eine Umstellung in der Ernährung (bei Insektenfressern durch Omnivorie [Omnivoren] – mit Früchteverzehr – zur Begünstigung der Fettdepotbildung), f) eine Reihe von hormonellen Veränderungen, vor allem Änderungen der Höhe von Hormonspiegeln in Bezug auf komplexe Wirkungsgefüge, g) die Ausbildung typischer Verhaltensweisen: bei Nachtziehern Reduktion der Tagaktivität, vor allem in der zweiten Tageshälfte, und Entwicklung von nächtlicher Zugaktivität (Aktivitätsbereitschaft); bei vielen Arten bilden sich Zuggemeinschaften, die im Verbandflug der Kräfteersparnis („Windschatteneffekt“) sowie dem Auffinden von Nahrungsquellen, Thermikzonen und Rastplätzen dienen, das Risiko gegenüber Beutegreifern mindern oder die Orientierungsleistung verbessern.
Steuerung des Zugs
Nur wenige Zugvogeltypen werden rein durch exogene Faktoren gesteuert, wie vor allem „Flüchter“ (Mangelflucht, speziell Winter-, Kälte-, Eis-, Hochwasser-, Feuerflucht u.a.). Bei den meisten Zugvögeln spielen endogene (genetische) Faktoren (endogene Ursachen) eine wichtige Rolle, bei ausgeprägten Zugvögeln sowohl für die Vorbereitungen für den Zug, seine Auslösung, den Ablauf als auch die Beendigung. Für mehr als 20 Zugvogelarten (aus 5 Kontinenten) ist gezeigt worden, daß sie über endogene Jahresperiodik (circannuale Rhythmik, biologische Jahreskalender; Chronobiologie [Farbtafel II], Jahreszyklen; vgl. Abb. 2 ) verfügen. Diese Körperrhythmen produzieren das gesamte Muster jahresperiodischer Vorgänge auf genetischer Grundlage ungefähr zeitgerecht, und die genaue Feineinstellung auf das biologische Kalenderjahr erfolgt hauptsächlich durch die Photoperiodizität – speziell die Änderungen der Tageslichtdauer im Jahresverlauf. Auf der Basis endogener Körperrhythmen haben Zugvögel eine ganze Reihe von genetisch gesteuerten Programmen entwickelt, die durch Selektion rasch verändert und angepaßt werden können (s.u.). Nach heutiger Vorstellung sind Vögel allgemein mit Genen für Ziehen und Nichtziehen ausgestattet. Ob ein Vogel Zugvogel (und kein Standvogel) ist, also „Zugtrieb“ besitzt, hängt davon ab, wie viele „Zuggene“ er geerbt hat. Ziehen ist somit ein quantitativ genetisches Merkmal ( vgl. Abb. 4 ). Erbfaktoren bestimmen auch den Aufbruch zum Zug, seinen Ablauf und die Beendigung. Das geschieht durch komplexe Raum-Zeit-Programme, die vor allem zugunerfahrene, erstmals und häufig allein wandernde Individuen auf verblüffende Weise „automatisch“ in ihnen unbekannte, aber ganz bestimmte geographische Regionen als Ruheziel („Winterquartier“) führen. Kuckucke z.B. können von nicht ziehenden Zaunkönigen, kurze Strecken wandernden Rotkehlchen oder extrem weit ziehenden Sumpfrohrsängern aufgezogen werden, die zudem in ganz verschiedene – westliche bzw. östliche – Richtungen wandern. Dennoch ziehen die nachts allein wandernden Jungkuckucke von Mitteleuropa einheitlich nach Süden in Ruheziele im südlichen Afrika, wo sie überwintern. Wie sie und viele andere Erstzieher das schaffen, erklärt die sog. Vektor-Navigations-Hypothese (s.u.).
Orientierungsmechanismen
Kreuzungsversuche mit Mönchsgrasmücken haben gezeigt, daß sie über angeborene, genetisch determinierte Zugrichtungen verfügen ( vgl. Abb. 5 ), und eine Fülle von Daten macht das für Zugvögel allgemein wahrscheinlich. Dabei kann genetische Determinierung auch Richtungsänderungen (Zugknicks, Bogenzug) mit einschließen. Weiterhin ist davon auszugehen, daß gutes Orientierungsvermögen nicht nur für Zugvögel typisch ist, sondern für Vögel generell. Auch nicht ziehende Arten zeigen nach Versetzungsversuchen hervorragende Heimkehrleistungen (Heimfindungsvermögen) – das beste Beispiel hierfür sind die Brieftauben. In Bezug auf die Orientierungsleistung sind 2 Kategorien zu unterscheiden: 1) die Richtungsorientierung oderKompaßorientierung (beim erstmals wandernden Jungvogel, Einhalten einer bestimmten, endogen vorgegebenen Richtung, die auch nach Versetzung oder Verdriftung beibehalten wird) und 2) die Zielorientierung oder Navigation (Zielfinden; beim erfahrenen Vogel bei Versetzung Rückkehr zu einem bekannten Ort – z.B. Taubenschlag – praktisch von jedem unbekannten Auflaßort aus). Die Mechanismen des Ziel-(Heim-)findens sind bislang (bis auf einen Spezialfall, s.u.) unbekannt. Man vermutet eine sog. Bikoordinaten-Navigation auf der Basis von 2 geophysikalischen Größen (z.B. des Erdmagnetfelds). Die Kompaßorientierung beruht, wie die Bezeichnung bereits sagt, auf der Anwendung von Kompassen. Für Vögel sind 3 sicher nachgewiesen: Beim Sonnenkompaß wird die Sonne als Richtgröße verwendet, wobei der Tagesgang der Sonne verrechnet werden muß. Dies geschieht mit Hilfe der circadianen, also endogenen Tagesperiodik (circadiane Rhythmik, innere Uhr), zudem kommt es zu Anpassungen an den Jahresgang der Sonne. Die entscheidende Größe für die Sonnenkompaßorientierung ist der Azimut (der Winkel auf dem Horizontkreis zwischen Meridian und Höhenkreis). Beim Magnetkompaß ist die Erde als riesiger Magnet Bezugsgröße, wobei Vögel die Neigungswinkel der Feldlinien zur Erdoberfläche (Magnetfeld [Abb.]) verwenden – daher auch Inklinationskompaß genannt. Der Sternenkompaß (Astrotaxis) beruht – wie bei der Verwendung durch den Menschen – auf der scheinbaren Rotation des Himmelsgewölbes um den (nördlichen bzw. südlichen) Himmelspol, in dessen unmittelbarer Nähe (auf der Nordhalbkugel) der Polarstern steht, der damit eine Referenz für Nord ergibt. Für die Südhalbkugel sind entsprechende „Referenzpunkte“ am Himmel anzunehmen. Weitere Richtgrößen bilden sehr wahrscheinlich der sog. Sonnenuntergangspunkt, das Polarisationsmuster des Himmelslichts (Polarisationskompaß), Infraschallmuster (von Gebirgen, Ozeanen usw.) sowie Duftfelder der Erdoberfläche (olfaktorische Orientierung; Geruchsorientierung). Über die Zielorientierung (Navigation) liegen rund 10 Hypothesen vor (u.a. das Karte-Kompaß-Konzept). Experimentell gut abgesichert ist bisher nur die Vektor-Navigations-Hypothese. Nach ihr finden erstmals und allein wandernde Zugvögel ihre zunächst unbekannten, aber geographisch definierten Ruheziele („Winterquartiere“) über einen Vektor, der sich zusammensetzt aus einer ererbten Zugrichtung und einem ebenfalls genetisch determinierten Zeitprogramm, das Zugvögel gerade so lange (Tage, Wochen oder Monate) zugaktiv sein läßt, wie zur Bewältigung der Wegzugstrecke erforderlich ist, so daß das Ruheziel im Winter „automatisch“ erreicht werden kann. Die sensorische Basis zur Perzeption von polarisiertem Licht (Polarisation; Polarisationssehen) und Infraschall ist experimentell gut bearbeitet. Die Wahrnehmung von Komponenten des Erdmagnetfelds (magnetischer Sinn) und von Spurengasen (zur olfaktorischen Orientierung) ist noch weitgehend unerforscht.
Ökologische Beziehungen
Obwohl der Vogelzug in hohem Maße durch Selektion (von genetisch determinierten Raum-Zeit-Programmen) den jeweiligen Umweltverhältnissen der Gebiete zwischen Brutplatz und Ruheziel angepaßt wird, ergeben sich dennoch sowohl auf dem Zug als auch im Ruheziel wechselnde ökologische Bedingungen, die unmittelbare Reaktionen verlangen. Nach jedem Aufbruch z.B. ist unklar, wie der nächste Rastplatz beschaffen sein wird. Bestehen Wahlmöglichkeiten im Hinblick auf Habitate mit unterschiedlichen Rastplatzbedingungen (z.B. in Bezug auf das Nahrungsangebot oder die Konkurrenz; Habitatselektion), kann die Auswahl optimiert werden durch (visuelle) Gestaltwahrnehmung, die auf angeborenen und erlernten Komponenten (Lernen) beruht. Auch meteorologische Ereignisse nehmen vielfach Einfluß auf das aktuelle Zuggeschehen. Treten z.B. während einer Zugetappe außergewöhnliche Wetterverhältnisse auf (vor allem Starkregen [Niederschlag], Sturm, Nebel), denen nicht in andere Höhenschichten ausgewichen werden kann, wird der Zug vielfach für Stunden oder einen Tag (selten länger) unterbrochen. Kommt es zu stärkeren Verdriftungen, nehmen Altvögel, die meist bestimmte Ziele anfliegen und nicht nur einer Kompaßrichtung folgen wie Jungvögel (s.o.), vielfach später eine Kurskorrektur vor. Relativ große Verhaltensvariabilität wird weiterhin vor allem auch durch unzureichende – nicht den saisonalen programmierten Sollwerten entsprechende – Fettdepots bewirkt. Zu geringe Fettdepots führen in Rastgebieten oft zu Hyperaktivität, die zu besseren Nahrungsquellen führen kann sowie zu längerer Verweildauer – bis zum Aufbruch zur nächsten Zugetappe. Gelangen Vögel mit zu geringen Fettdepots in den Bereich ökologischer Barrieren wie der Sahara, setzen sie vielfach weder zu einem Non-Stop-Flug noch zur Rast mitten in der Wüste an, sondern suchen gezielt Oasen auf, die sie auch aus großer Höhe erkennen, um dort wieder Fettdepots zu bilden. Erreichen relativ magere Vögel offenes Wasser, fliegen sie zum Teil wieder zurück (sog. Umkehrzug), um zunächst im Landbereich die Fettdepots zu vergrößern. Ist das Ruheziel bereits erreicht, und verschlechtern sich die Bedingungen dort, kann – sofern die Winter-Mauser noch nicht begonnen hat – noch einmal weitergewandert werden (fakultativer Zug, z.B. Trockenflucht bei Dürre), bis besser geeignete Ziele erreicht werden.
Gefährdung und Schutz von Zugvögeln
Vögel sind derzeit durch vielerlei menschliche Aktivitäten (vor allem Habitatzerstörungen, exzessive Landwirtschaftspraktiken, Tourismus, direkte Verfolgung) weltweit gefährdet, was sich im Bestandsrückgang vieler Arten bemerkbar macht. Zugvögel sind von Bestandsrückgängen besonders betroffen (Rote Liste), da sie – im Gegensatz zu Standvögeln – nicht nur in 1, sondern in 3 Gebieten Gefahren ausgesetzt sind: dem Brutgebiet, den Rastgebieten (Zwischenzielen) sowie dem Ruheziel. In Mitteleuropa beträgt der Rückgang der Langstrecken ziehenden Kleinvögel derzeit jährlich rund 1 Prozent, wobei manche Arten geringer (z.B. Gartengrasmücke, Grauschnäpper, Teichrohrsänger), andere stärker betroffen sind (z.B. Dorngrasmücke, Fitislaubsänger, Gartenrotschwanz). Leider sind Gegenmaßnahmen nur begrenzt möglich und nur in sehr geringem Maße wirksam, obwohl sich die verschiedensten Einrichtungen darum bemühen, so z.B. UNESCO, UNEP (United Nations Environment Programme), WWF (Worldwide Fund for Nature), ICB (International Council for Bird Preservation), IUCN (International Union for Conservation of Nature), die „Bonner Konvention zur Erhaltung der wandernden wildlebenden Tierarten“ (Bonner Konvention), CIC (Conseil International de la Chasse) und viele andere (Birdlife International, Fauna-Flora-Habitat-Richtlinie, Important Bird Area, Naturschutz, Vogelschutzrichtlinie). 3 Hauptgründe sind für das letztendliche Scheitern von Maßnahmen verantwortlich: 1. sind häufig auch bei schon länger anhaltenden Bestandsrückgängen trotz intensiver Forschung die Ursachen dafür nicht klar zu benennen, weil sie sehr komplex sind und zudem variabel auftreten können (wie z.B. beim Weißstorch); 2. lassen sich häufig, wenn Ursachen erkannt worden sind, Gegenmaßnahmen aus technischen Gründen – wie z.B. im Falle der Sahel-Dürre, die viele Zugvogelarten schädigt – oder aus politischen Gründen – wie im Fall der derzeitigen vogelfeindlichen Landwirtschaftspraxis in Mitteleuropa – kaum durchführen; 3. fehlt es häufig in der Gesellschaft an der Bereitschaft, Zugvogelschutz (Vogelschutz) Priorität einzuräumen (z.B. bei der Schaffung von Ruhezonen unter Verzicht auf Freizeit- oder Tourismuseinrichtungen). Ein Sonderweg im Zugvogelschutz wurde beim Weißstorch begangen. Als die westeuropäischen Populationen zum Teil ausgestorben waren (Belgien 1895, Schweiz 1950, Schweden 1955, Niederlande 1991), wurden in speziellen Brutstationen Störche gezüchtet und anschließend ausgebürgert. Diese semidomestizierten Vögel haben inzwischen größere Populationen aufgebaut, bedürfen jedoch regelmäßiger Zufütterung – vor allem die großenteils nicht mehr wegziehenden Altvögel –, so daß die Maßnahme umstritten ist. Immerhin hat sie gebietsweise die Storchenpopulation vor dem völligen Erlöschen bewahrt; bei den meisten Zugvogelarten wären jedoch derartige Zucht- und Ausbürgerungsverfahren nicht durchführbar.
Vogelzug und rezente Klimaerwärmung
Obwohl Vogelzug in hohem Maße genetisch gesteuert wird und Zugcharakteristika relativ hohe Erblichkeit (Heritabilität) aufweisen, sind Zugvögel in ihrem Verhalten keineswegs starr. Sie können sich vielmehr erstaunlich schnell an neuartige Umweltbedingungen anpassen. Das kann theoretisch auf zweierlei Weise geschehen: 1. durch phänotypische Plastizität (unmittelbare individuelle Anpassung) oder 2. durch Selektion (erhöhte Fitness [Adaptationswert, inclusive fitness], höchster Reproduktionserfolg [Fortpflanzungserfolg] der Individuen mit der günstigsten Genkombination). Da die Kleinvögel höherer Breiten in der Regel nur etwa eineinhalb Jahre alt werden, können sie nahezu keine Erfahrungen über Jahre hinweg sammeln, die sie dann gezielt anwenden könnten. Zudem setzen die ererbten Raum-Zeit-Programme enge Grenzen für individuelle Plastizität. Damit ist die 2. Anpassungsmöglichkeit – über Selektion – wahrscheinlicher. Sie ist sowohl im Experiment (Zweiweg-Selektionsexperiment mit Teilziehern [ vgl. Abb. 6 ], schnelle Zucht von Zug- und Standvögeln, s.o.) als auch im Freiland nachgewiesen. Mönchsgrasmücken, die zum Überwintern seit den 1960er Jahren von Mitteleuropa zunehmend auf die Britischen Inseln anstatt in den Mittelmeerraum (Mediterranregion) wandern, haben ihre neue Zugrichtung (West-Nord-West statt Süd-West) durch Selektion etabliert. Voraussetzung für eine derartig schnelle Selektion ist neben einer relativ hohen Heritabilität eine ausreichend große genetische Variabilität, die bei verschiedenen untersuchten Zugparametern gegeben ist. Die globale Klimaerwärmung (Klimaänderungen) bewirkt bei Zugvögeln derzeit vor allem systematische Veränderungen in 6facher Hinsicht: 1) Abnahme des Zugumfangs (vermehrte Überwinterung im Brutgebiet), 2) Verkürzung von Zugstrecken (Überwinterung näher zum Brutgebiet), 3) Verspätung des Wegzugs, 4) Verfrühung des Heimzugs, 5) Änderung von Zugrichtungen in neuartige, nähergelegene „Winterquartiere“ (wie im oben geschilderten Fall der Mönchsgrasmücke), 6) Ausdehnung von Brutgebieten in höhere Breiten (derzeit auch in Europa bei vielen Arten, besonders augenfällig bei verschiedenen weißen Reiherarten [Reiher], Bienenfressern u.a.). Eine anhaltende starke Klimaerwärmung dürfte die weitere Entwicklung vom Zug- zum Standvogelverhalten beschleunigen. In höheren Breiten könnte der Zug sogar weitgehend verschwinden, in äquatornäheren Trockengebieten (durch Trockenflucht) eher mehr aufleben. Insgesamt sind größere Umstrukturierungen kontinentweiter Avifaunen zu erwarten, die bereits im Gange sind. Audubon (J.J.), Ethologie, Ethologie (Geschichte der), Heinroth (J.Ch.A.), kognitive Landkarte, Lichtverschmutzung, Menotaxis, Migration, Mobilität, Ortsprägung, Reichenow (A.), Synchronisation, Thienemann (J.).
Lit.:Berthold, P.: Vogelzug. Eine aktuelle Gesamtübersicht. Darmstadt 42000. Berthold, P.: Control of Bird Migration. London 1996. Dingle, H.: Migration. The Biology of Life on the Move. Oxford 1996. Perrin, J.: Nomaden der Lüfte. Hildesheim 2002. Wiltschko, R., Wiltschko, W.: Magnetic Orientation in Animals. Berlin 1995.
;)
Vogelzug
Abb. 1: Eine Mönchsgrasmücke (Sylvia atricapilla) bei der „Produktion“ von Zugunruhe (Zugaktivität im Käfig, einer Art „Ziehen im Sitzen“; Videoaufnahme der nächtlichen Aktivität dieser nachts ziehenden Art bei Infrarotlichtbeleuchtung).
;)
Vogelzug
Abb. 2: Endogene Jahresperiodik (circannuale Rhythmik) der Zugunruhe (gemessen als nächtliche lokomotorische Aktivität, „Hüpfer“), des Körpergewichts und der Mauser einer männlichen Gartengrasmücke (Sylvia borin). Der handaufgezogene Vogel, Ende Mai geschlüpft, wurde im Juni in konstante Bedingungen (ein Licht-Dunkel-Verhältnis von 10:14 Stunden) übergeführt und dort 10 Jahre lang gehalten; dargestellt sind Daten aus den ersten 3 Versuchsjahren. HZ Heimzugperiode, WZ Wegzugperiode.
;)
Vogelzug
Abb. 3: Beispiele für Wanderrouten von Langstreckenziehern auf dem Wegzug: 1 Wanderregenpfeifer (Pluvialis dominica) und andere Watvögel von Alaska zu Inselgruppen im Pazifik, 2 Präriebussard (Buteo swainsoni), 3 Transgolfzug vieler nordamerikanischer Arten, 4 Kappenwaldsänger (Dendroica striata, Transatlantikzug), 5 Knutt (Calidris canutus rufa), 6 Küstenseeschwalbe (Sterna paradisaea), 7 Kampfläufer (Philomachus pugnax), 8 Transsaharazug vieler eurasischer Arten, 9 Steinschmätzer (Oenanthe oenanthe, Alaska-Population), 10 Rauchschwalbe (Hirundo rustica, 3 verschiedene Populationen in unterschiedliche Winterquartiere), 11 Amurfalke (Falco amurensis, dargestellt ist nur die Ozeanüberquerung), 12 Knutt (Calidris canutus rogersi), 13 zirkulärer Weg- und Heimzug des Kurzschwanz-Sturmtauchers (Puffinus tenuirostris).
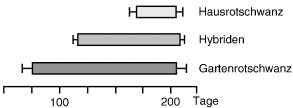
Vogelzug
Abb. 4: Die Zugaktivität von Gartenrotschwänzen (frühzeitig und lang) und Hausrotschwänzen (spät und kurz) und deren Hybriden, die sich intermediär verhalten, womit die genetische Steuerung der Zugaktivität belegt ist.
;)
Vogelzug
Abb. 5: Zugrichtungspräferenzen von Mönchsgrasmücken (Sylvia atricapilla), die in Orientierungskäfigen ermittelt wurden. Innerer Kreis, links: süddeutsche Vögel, die in freier Natur nach Südwesten in den Mittelmeerraum ziehen; rechts: Vögel aus Ostösterreich, die nach Südosten abwandern; äußerer Kreis: Hybriden beider Populationen, die sich intermediär verhalten. Kleine Pfeile: Werte von Einzelvögeln, große Pfeile: Mittelwerte.
;)
Vogelzug
Abb. 6: Ergebnisse eines Zweiweg-Selektionsexperiments mit (über 700) teilziehenden Mönchsgrasmücken (Sylvia atricapilla) aus Südfrankreich, bei den Nichtziehern bis zur F6-Generation, bei den Ziehern bis zur F3-Generation. 267: Anzahl der handaufgezogenen Vögel der Ausgangspopulation, die aus rund 75% Zugvögeln und 25% Standvögeln besteht.





Wenn Sie inhaltliche Anmerkungen zu diesem Artikel haben, können Sie die Redaktion per E-Mail informieren. Wir lesen Ihre Zuschrift, bitten jedoch um Verständnis, dass wir nicht jede beantworten können.